Brain. 2013 Jan 31.
Politis M, Loane C, Wu K, O’Sullivan SS, Woodhead Z, Kiferle L, Lawrence AD, Lees AJ, Piccini P.
Source: Division of Brain Sciences, Department of Medicine, Hammersmith Hospital, Imperial College London, London W12 0NN, UK.
Abstract
Hypersexuality with compulsive sexual behaviour is a significant source of morbidity for patients with Parkinson’s disease receiving dopamine replacement therapies. We know relatively little about the pathophysiology of hypersexuality in Parkinson’s disease, and it is unknown how visual sexual stimuli, similar to the portrayals of sexuality in the mainstream mass media may affect the brain and behaviour in such susceptible individuals.Here, we have studied a group of 12 patients with Parkinson’s disease with hypersexuality using a functional magnetic resonance imaging block design exposing participants to both sexual, other reward-related and neutral visual cues. We hypothesized that exposure to visual sexual cues would trigger increased sexual desire in patients with Parkinson’s disease with hypersexuality that would correspond to changes in brain activity in regions linked to dopaminergically stimulated sexual motivation.
Patients with Parkinson’s disease with hypersexuality were scanned ON and OFF dopamine drugs, and their results were compared with a group of 12 Parkinson’s disease control patients without hypersexuality or other impulse control disorders. Exposure to sexual cues significantly increased sexual desire and hedonic responses in the Parkinson’s disease hypersexuality group compared with the Parkinson’s disease control patients. These behavioural changes corresponded to significant blood oxygen level-dependent signal changes in regions within limbic, paralimbic, temporal, occipital, somatosensory and prefrontal cortices that correspond to emotional, cognitive, autonomic, visual and motivational processes.
The functional imaging data showed that the hypersexuality patients’ increased sexual desire correlated with enhanced activations in the ventral striatum, and cingulate and orbitofrontal cortices. When the patients with Parkinson’s disease with hypersexuality were OFF medication, the functional imaging data showed decreases in activation during the presentation of sexual cues relative to rest.
These deactivations were not observed when the patients were ON medication, suggesting that dopamine drugs may release inhibition within local neuronal circuits in the cerebral cortex that may contribute to compulsive sexual behaviour.
The findings of this study have implications with respect to the potential influence of cue exposure via exposure to mass media in enhancing libido, which in this group of vulnerable patients can lead to devastating social consequences and occasionally, custodial sentences.
Stimulation through exposure to sexual visual cues in patients with Parkinson’s disease with hypersexuality provides a motivational impetus for seeking this reward behaviour through activations and deactivations of cerebral cortex.
Introduction
Impulse control disorders including hypersexuality and compulsive sexual behaviour are a significant source of morbidity for patients with Parkinson’s disease receiving dopamine replacement therapy (Weintraub et al., 2006; Evans et al., 2009; Voon et al., 2009). Hypersexuality with compulsive sexual behaviour commonly manifests with excessive requests for sex from a spouse or a partner, increased pornographic interest, compulsive masturbation, promiscuity with visits to prostitutes, and in some premorbidly predisposed individuals, to paraphillias (e.g. sadism, bestiality, transvestitism; Quinn et al., 1983; Voon et al., 2006). The prevalence of hypersexuality in Parkinson’s disease was ∼3.5% in a large study of patients receiving dopamine agonist therapy with males being more likely than females to have diagnosed hypersexuality (Weintraub et al., 2010). A newer but smaller study has suggested a somewhat higher prevalence of hypersexuality in Parkinson’s disease of 7% and a link with l-DOPA (Hassan et al., 2011). Earlier studies have also shown a connection between l-DOPA and hypersexuality (Ballivet et al., 1973; Brown et al., 1978; Uitti et al., 1989).
Sexual content in the media has become more explicit and accessible to the general public, and pornography is readily available on the internet. It has been claimed that continuous exposure to sexual material is able to stimulate excessive socially unacceptable preoccupations with sex in some constitutionally vulnerable individuals and individuals taking dopaminergic drugs (Rees et al., 2007). Studies in monkeys indicate that a synergistic interaction between dopamine drugs and sexual cues markedly enhances excessive sexual activity (Pomerantz, 1990). The importance of cues being highly associated with reward is consistent with incentive salience theory, as ‘wanting’ is produced by a synergistic interaction between the current state of the mesolimbic dopamine system and the presence of rewards or their cues (Zhang et al., 2009; Berridge, 2012).
PET (Redouté et al., 2000) and functional MRI (Arnow et al., 2002; Hamann et al., 2004; Walter et al., 2008) studies of the neural processing of visual sexual stimuli in healthy individuals have shown that a number of regions including the ventral striatum, hypothalamus, amygdala, cingulate and orbitofrontal cortex are implicated in visual processing of sexual stimuli. In a recent study, using PET and indirect measures of striatal dopamine release, it was shown that patients with a Parkinson’s disease impulse control disorder had greater ventral striatum dopamine release following reward-related visual cue exposure compared with a group of Parkinson’s disease control patients (O’Sullivan et al., 2011). This finding was in line with the incentive sensitization theory, which proposes that compulsive reward seeking arises from excessive attribution of incentive salience (or ‘wanting’) for rewards and their cues, as a result of progressive neuroadaptations in dopamine projections to ventral striatum-related motivation circuitry (Berridge et al., 2009).
Experimental studies have shown that the dopaminergic neural circuitries in areas such as the ventral striatum, hypothalamus, amygdala and the medial prefrontal cortex play a role in sexual motivation and pursuit, especially in response to sexual cues (Pfaus, 2010; Stolzenberg and Numan, 2011). Sensitization by dopaminergic drugs has been shown to enhance pursuit of even natural rewards, including sexual incentives (Fiorino and Phillips, 1999; Nocjar and Panksepp, 2002; Afonso et al., 2009), as a result of excessive incentive salience attribution to reward cues. Moreover, sexual behaviour and dopaminergic drugs coactivate a population of neurons in the ventral striatum, amygdala, hypothalamus and anterior cingulate cortex, identifying potential sites where drugs can influence sexual behaviour (Frohmader et al., 2010; Holder et al., 2010).
In line with the incentive sensitization theory, hypersexuality in Parkinson’s disease could be associated with increased processing in brain regions linked to sexual motivation and cue reactivity, and these activations could be potentiated by dopaminergic medication. In the present study, we hypothesized that patients with Parkinson’s disease with hypersexuality would show aberrant sexual cue related activity in these regions linked to sexual motivation, which would be modulated by dopaminergic drugs, and linked to enhanced sexual motivation. We sought to investigate this by examining blood oxygen level-dependent (BOLD) signal alterations with functional MRI (by comparing responses in an ON medication scan with an OFF medication scan) and correlate the imaging results with behavioural assessments of sexual motivation.
Patients and methods
Participants and clinical characteristics
Twenty-four non-demented patients with idiopathic Parkinson’s disease were studied (Tables 1 and 2). Twelve of these fulfilled the proposed operational diagnostic criteria for hypersexuality (Voon et al., 2006; Supplementary Table 1). The other 12 patients with Parkinson’s disease had no history of hypersexuality or other impulse control disorders and served as the control group. Frohmader et al. (2011) have shown that the effects of dopaminergic drugs on compulsive sexual behaviour in animal models are dependent on the concurrent experience of dopaminergic drugs and sexual experience. Similarly, drug-induced hypersexuality in Parkinson’s disease only develops in the context of repeated medication use and does not occur acutely in de novo patients (Giladi et al., 2007). Hence, we compared patients with Parkinson’s disease with hypersexuality to a control group of medicated patients with Parkinson’s disease.
Participants’ clinical characteristics
| Clinical characteristics | Parkinson’s disease with hypersexuality | Parkinson’s disease control patients | P-value |
|---|---|---|---|
| Number of subjects | 12 | 12 | |
| Age (years ± SD) | 55.2 ± 9.2 | 62.3 ± 9.7 | 0.077 b |
| Sex | 11 M/1 F | 10 M/2 F | |
| Disease duration (years ± SD) | 9.6 ± 5.2 | 10.1 ± 6.4 | 0.85b |
| UPDRS OFF motor (Part III) score (mean ± SD)a | 40.2 ± 10.1 | 34.9 ± 9.9 | 0.21b |
| UPDRS ON motor (Part III) score and % improvement (mean ± SD)a | 23.1 ± 8.2 (43.8 ± 9.7%) | 20.0 ± 5.5 (41.4 ± 11.7%) | 0.29 (0.59)b |
| Mini-Mental State Examination (mean ± SD) | 29.8 ± 0.4 | 28.9 ± 2.2 | 0.30c |
| Daily LEDTOTAL (mg ± SD) | 600 ± 327 | 778 ± 278 | 0.17b |
| Daily LEDL-DOPA (mg ± SD) | 288 ± 326 | 646 ± 264 | <0.05c |
| Daily LEDDA (mg ± SD) | 311 ± 183 | 132 ± 143 | <0.01c |
a The average of five evaluations on five different days.
b Unpaired t-test.
c Mann–Whitney test.
Parkinson’s disease control patients = patients with Parkinson’s disease without hypersexuality or other impulsive-compulsive behaviours; M = male; F = female; SD = standard deviation; UPDRS = Unified Parkinson’s Disease Rating Scale; LED = levodopa equivalent. The dose is calculated similarly to previous report (Politis et al., 2010): LED (mg) = (1 × = levodopa) + (0.77 × levodopa CR) + (1.43 × levodopa + entacapone) + (1.11 × levodopa CR + entacapone) + (20 × ropinirole) + (20 × ropinirole ER) + (100 × pramipexole) + (30 × rotigotine) + (10 × bromocriptine) + (8 × apomorphine) + (100 × pergolide) + (67 × cabergoline) LED formula, in levodopa/carbidopa or benserazide hydrochloride: only levodopa is calculated.
Patients with Parkinson’s disease with hypersexuality
| Subjects | Sex | Age | Type of hypersexuality behaviour | Other ICDs | Dopaminergic therapy | Daily LEDDA | Daily LEDL-DOPA |
|---|---|---|---|---|---|---|---|
| HS1 | M | 46 | Pornography/repetitively engaging in sexual fantasies and urges | CS | Cabergoline | 280 | 0 |
| HS2 | M | 65 | Sex with people with whom he normally would not associate/spends excessive time obsessing about sex or engaged in sexual activity | Pramipexole | 267 | 0 | |
| HS3 | M | 72 | Sex with people with whom he normally would not associate | Ropinerole, levodopa | 180 | 700 | |
| HS4 | M | 65 | Empty affairs/involvement with internet pornography | Pramipexole | 200 | 0 | |
| HS5 | M | 50 | Patient refused to disclose | BE, CS | Ropinerole, levodopa | 180 | 800 |
| HS6 | F | 55 | Increased sexual activity | CS | Pramipexole | 200 | 0 |
| HS7 | M | 53 | Pornography | Pramipexole, levodopa | 240 | 260 | |
| HS8 | M | 53 | Empty affairs/obsessive sexual thoughts | PG, BE, CS | Ropinerole | 360 | 0 |
| HS9 | M | 60 | Patient refused to disclose | BE, CS | Ropinerole, levodopa | 300 | 600 |
| HS10 | M | 41 | Pornography/frequent visits to prostitutes | DDS | Cabergoline, levodopa | 530 | 500 |
| HS11 | M | 45 | Video pornography/involvement with internet pornography | DDS | Pramipexole, levodopa | 200 | 600 |
| HS12 | M | 57 | Involvement with internet pornography | DDS, PG, BE | Ropinerole | 800 | 0 |
BE = binge eating; CS = compulsive shopping; DA = dopamine agonist; DDS = dopamine dysregulation syndrome; HS = hypesexuality; ICD = impulse control disorder; PG = pathological gambling.
The clinical assessment battery included Hoehn and Yahr staging, the motor part (Part III) of the Unified Parkinson’s Disease Rating Scale (UPDRS), Mini-Mental State Examination and calculation of daily l-DOPA equivalent dose (LED). Eight of 12 patients with Parkinson’s disease hypersexuality exhibited at least one additional impulse control disorder (Tables 1 and 2).
The study received ethical approval from the Hammersmith and Queen Charlotte’s Hospitals Research Ethics Committee. Written informed consent was obtained from all participants in accordance with the Declaration of Helsinki.
Behavioural assessments
Before and after scanning, participants were asked to rate their libido and sexual desire on a visual analogue scale (10 cm), anchored between ‘the lowest ever’ (0 cm = 0 points) and ‘the most ever’ (10 cm = 10 points). Both scales were self-reported and related to how the participants felt within the past hour only. Single-item desire scales have been shown to be as reliable as longer multi-item questionnaires (West and Ussher, 2010). As most of the patients with Parkinson’s disease hypersexuality had additional impulse control disorders, similar self-reported desire scales for gambling, drugs and food were also administered.
Functional magnetic resonance imaging scanning procedures
Participants were scanned on two separate mornings (between 11:00 and 13:00 h) 7 days apart in a cross-over randomized design after skipping breakfast and stopping medication at least 18 h before scanning. Participants were scanned in the practically defined OFF medication condition in one scan and in the ON medication condition after receiving an oral dose of l-DOPA/benserazide (200/50 mg) dispersible 45 min before the scan starting. Motor performance was assessed with the UPDRS motor scores at baseline and immediately before scanning to ensure the patient had responded to the medication (defined as >25% improvements in UPDRS-III motor scores). The use of l-DOPA was chosen because all patients with Parkinson’s disease were previously taking this, whereas not all were on the same dopamine agonist. Moreover, l-DOPA can increase sexual motivation in Parkinson’s disease, and hypersexuality behaviour has been observed in combination with concurrent l-DOPA treatment, not just dopamine agonist treatment (Ballivet et al., 1973; Brown et al., 1978; Hassan et al., 2011).
Participants were placed in the scanner with headphones and padding round the head, the shoulders and arms to ensure as little area for movement as possible. Movements were monitored throughout the scans and in the case of excessive movements, the scan was either restarted or the relevant volumes were removed from the analysis (5.2% of runs were restarted owing to tremor or dyskinesias). Imaging acquisition was conducted on a 3 T Philips Intera whole-body scanner. Whole-brain data were acquired with 199 volumes of T2*-weighted gradient-echo echo-planar imaging in ascending order with an automated higher-order shim procedure (slice thickness 3.25 mm; repetition time 3000 ms; echo time 30 ms; 90° flip angle; field of view 190 × 219; matrix 112 × 112). The slice acquisition angle was set at −30° from the anterior–posterior commissure line to reduce frontal lobe signal dropout owing to the air sinuses, with a z-shim gradient correction to compensate for through-plane susceptibility gradients (Deichmann et al., 2003; Goldstone et al., 2009). One high-resolution T1-weighted turbo field echo structural scan was also collected (echo time 4.6 ms; repetition time 9.7 ms; 8° flip angle; field of view 240 mm).
Functional magnetic resonance imaging paradigm
During the functional MRI scan, five types of colour images were presented in a block design: (i) dopaminergic drugs cues; (ii) appetizing food cues; (iii) money and gambling cues; (iv) sexual cues; and (v) neutral cues. The vast majority of reward-related and neutral visual cues were obtained from the International Affective Picture System (Lang et al., 2008) and were supplemented by freely available images obtained from websites. A visual sexual cue was defined as including depictions of intimate touching, passionate kissing, physical flirting and provocative images of women or men (gender based: males saw females and females saw males) with their bodies partly clothed. Neutral visual cues included landscapes and nature scenes, household objects and random patterns. The International Affective Picture System sexual visual stimuli have been previously validated and have been shown to induce significant levels of sexual arousal in previous functional MRI/psychophysiological studies (Bradley et al., 2001; Conaglen and Evans, 2006; Walter et al., 2008).
Images of similar resolution were presented in 14.7 s blocks in two runs lasting 9 min 56 s each. Each block contained seven different images from the same category, with a total of six blocks of each type shown in a pseudorandom block order with a randomized image order in each block. Run order was counterbalanced across participants and visits. Each image was displayed for 2100 ms, and each thematic block was followed by a 4000 ms rating slide whereby the participant had to rate from 1 to 5 how much they liked the block of images they saw (with 1 being ‘I hated it’ and 5 being ‘I loved it’). These ratings were made by voice, and they were recorded in a computer. The rating slide was followed by a 1000 ms interstimulus interval of a fixation cross. Images were viewed via a mirror mounted above an eight-channel RF head coil, which displayed images from a projector using the IFIS-SA image presentation system (In Vivo) and E-Prime software (Psychology Software Tools Inc).
Functional magnetic resonance imaging data analysis
Imaging data were analysed using Statistical Parametric Mapping version 5 (Wellcome Department of Imaging Neuroscience, UCL, UK). The first five volumes of each functional MRI run were discarded to allow for equilibrium effects, and all functional scans were realigned to the first scan of the run and then again to the mean of all of the volumes for motion and slice-timing correction. All scans included in the final analysis consisted of <2 mm of movement in each direction. We explored the data for residual artefacts using the TSDiffAna utility implemented in Statistical Parametric Mapping version 5, which produces mean and variance images for each functional volume (http://imaging.mrc-cbu.cam.ac.uk/imaging/DataDiagnostics). An artefact is defined as the co-occurrence of a variance spike and a mean intensity drop uncorrelated with experimental design. We did not detect any artefacts that needed more detailed inspection. The mean functional image was co-registered to the T1 structural image. The echo-planar images were then normalized into Montreal Neurological Institute standard stereotactic space using parameters from the segmented T1 structural image and smoothed using an 8 mm full-width at half-maximum Gaussian filter.
First-level analysis was performed at the single participant level where individual contrasts were created in a general linear model for each reward condition minus baseline (neutral pictures) and for sex minus other rewards (sex versus other rewards). The general linear model included six motion and nuisance parameters, accounting for movement and other nuisance artefacts occurring in three directions of translation (displacement) and three axes of rotation (x, y, z translation and x, y, z rotation) for each run. Contrasts of interest produced at this stage (e.g. sex versus neutral) were used in the second-level, group random effects analysis. A statistical threshold of P < 0.001 uncorrected and cluster extent over 10 voxels (2 × 2 × 2 mm) was used for activation using whole-brain analysis with correction for multiple comparisons using the false discovery rate at P < 0.05 (Genovese et al., 2002). Coordinates of peak voxel activation within the brain were determined at the group level for ON and OFF scans in patients with Parkinson’s disease with and without hypersexuality. We also performed an additional between-group (Parkinson’s disease control patients versus Parkinson’s disease with hypersexuality) and between-condition (OFF versus ON medication) analysis to investigate whether motion parameters were larger in one patient or condition group than the other and found that movement did not vary between groups and conditions (P > 0.1 in both cases).
As a regional a priori hypothesis existed, additional second-level random effects analyses (sex versus neutral and sex versus other rewards contrasts in ON and OFF medication conditions) were carried out for specific regions of interest using MarsBar (Brett et al., 2002) with the same statistical threshold (false discovery rate at P < 0.05). These regions were the orbitofrontal cortex, anterior cingulate cortex, posterior cingulate cortex, amygdala, ventral striatum and the hypothalamus. The models of orbitofrontal cortex, anterior cingulate cortex, posterior cingulate cortex and amygdala were taken from the Automated Anatomic Labelling library that accompanies MarsBar. The ventral striatum and hypothalamus object maps were constructed by drawing these regions of interest in ANALYZE medical imaging software (version 8.1, Mayo Foundation). These object maps were then used to mask out the rest of the brain allowing comparisons within specific volumes (1850 mm3 in each hemisphere for ventral striatum and 1380 mm3 in each hemisphere for the hypothalamus). Individual participants’ contrast (e.g. sex versus neutral) values of each region of interest for both ON and OFF medication scans were extracted for correlation with the behavioural data.
Statistical analysis
Statistical analyses were performed using SPSS (version 16, SPSS Inc) for Macintosh. Comparisons within groups (e.g. Parkinson’s disease with hypersexuality OFF scan sex versus neutral) were performed using paired t-tests and comparisons between groups (e.g. Parkinson’s disease with hypersexuality OFF sexual versus neutral—versus—Parkinson’s disease control patients sexual versus neutral) were performed using two sample t-tests. For the clinical and behavioural analysis, variance homogeneity and Gaussianity were tested with Bartlett and Kolmogorov–Smirnov tests. Parametric and non-parametric tests were used appropriately. Pearson correlation coefficient r and Spearman’s rho (ρ) (when variables were not normally distributed) were used to examine the relationship between individual sexual desire scores after exposure to sexual visual cues and individual bilateral BOLD region of interest contrast values for sexual versus neutral contrast in the ON and OFF medication states. The Bonferroni correction was used to counteract the problem of multiple comparisons.
Results
Clinical and behavioural analysis
Patients with Parkinson’s disease with hypersexuality were taking significantly more dopamine agonists and significantly less l-DOPA compared with Parkinson’s disease control patients. The two groups did not statistically differ in any of the other clinical characteristics (age, sex, disease duration, UPDRS part III OFF, ON and response after l-DOPA, Mini-Mental State Examination, daily LEDTOTAL) (Tables 1 and 2).
Before the OFF and the ON scan, patients with Parkinson’s disease with hypersexuality and Parkinson’s disease control patients did not differ on sexual desire scores (Table 3). After the OFF and the ON scan where participants had been exposed to sexual cues, patients with Parkinson’s disease with hypersexuality showed a significant increase in their sexual desire compared with their rating before the scan and also showed a significant increase compared with the sexual desire of the Parkinson’s disease control patients, the latter remaining relatively stable pre and post scan. Patients with Parkinson’s disease with hypersexuality had higher increases in their sexual desire in the ON compared with the OFF scan (Table 3). The Parkinson’s disease with hypersexuality and Parkinson’s disease control groups did not show any within or between group differences in food, gambling and drug desire before and after the ON and OFF scans (data not shown).
Sexual desire before and after exposure to sexual visual stimuli
| Before sexual visual stimuli | After sexual visual stimuli | P-value | |
|---|---|---|---|
| OFF scan | |||
| PD control patients (mean ± SE) | 1.98 ± 0.59 | 2.25 ± 0.45 | 0.31a |
| PD HS (mean ± SE) | 2.67 ± 0.56 | 3.70 ± 0.50 | <0.01a |
| P-value | 0.40b | <0.05b | |
| ON scan | |||
| PD control patients (mean ± SE) | 1.32 ± 0.28 | 2.12 ± 0.60 | 0.18d |
| PD HS (mean ± SE) | 2.01 ± 0.39 | 5.24 ± 0.41 | <0.001a |
| P-value | 0.15c | <0.001b | |
| OFF scan | ON scan | ||
|---|---|---|---|
| After-before sexual visual stimuli | |||
| PD control patients (mean ± SE) | 0.28 ± 0.26 | 0.80 ± 0.52 | 0.34d |
| PD HS (mean ± SE) | 1.04 ± 0.32 | 3.23 ± 0.51 | <0.01a |
| P-value | 0.08b | <0.01b | |
a Paired t-test.
b Unpaired t-test.
c Mann–Whitney test.
d Wilcoxon matched pairs test.
HS = hypersexuality; PD = Parkinson’s disease.
Patients with Parkinson’s disease with hypersexuality liked the sexual visual cues presented during the functional MRI significantly more than Parkinson’s disease control patients (quality of data = 85.2%; 3.4 ± 1.2 versus 2.1 ± 0.6, mean ± SD; P < 0.05, unpaired t-test with Welch correction), but there was no difference in their ratings for reward cues related to drugs, money and gambling, or food (P > 0.1 in all cases, unpaired t-test with Welch corrections). No differences were found between ON and OFF scans.
Region of interest functional magnetic resonance imaging analysis
The region of interest analysis showed that the patients with Parkinson’s disease with hypersexuality had stronger activity (increased BOLD signal) than the Parkinson’s disease control patients during exposure to sexual versus neutral visual cues in the OFF medication states in the following regions: orbitofrontal cortex (left: P < 0.001, right: P < 0.005), anterior cingulate cortex (left: P < 0.005, right: P < 0.001), posterior cingulate cortex (left: P < 0.001, right: P < 0.001), left amygdala (P < 0.05), ventral striatum (left: P < 0.05, right: P < 0.05) and hypothalamus (left: P < 0.005, right: P < 0.01) (Fig. 1C–H). For the same contrast in the ON medication state, a similar set of regional brain activations was observed, and there were no significant differences in activations between ON and OFF scans. When we compared the effect of OFF versus ON medication state just in the group of patients with Parkinson’s disease with hypersexuality, we found no differences in activation.
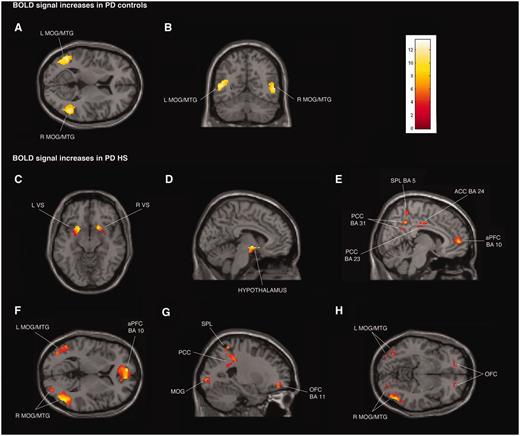
{kind=link}
Transverse, coronal and sagittal sections of statistical parametric maps showing significant BOLD signal increases (yellow–red areas) associated with exposure to common sexual imagery in 12 Parkinson’s disease (PD) control patients (A and B) and in 12 patients with Parkinson’s disease with hypersexuality (PD HS) (C–H) during ON and OFF medication states. BOLD signal increases are illustrated in (A and B) left and right middle temporal gyrus (MTG) and middle occipital gyrus (MOG) (x = −48, y = −59, z = 9), (C) ventral striatum (VS) (x = 18, y = 15, z = − 11), (D) hypothalamus (x = −5, y = −4, z = −9), (E) anterior prefrontal cortex (aPFC), anterior cingulate cortex (ACC), superior parietal lobule (SPL) and posterior cingulate cortex (PCC) (x = 8, y = −16, z = 33), (F) left and right middle temporal gyrus and middle occipital gyrus and anterior prefrontal cortex (x = −8, y = 56, z = 4), and (G and H) orbitofrontal cortex (OFC), superior parietal lobule, posterior cingulate cortex, and left and right middle temporal gyrus and middle occipital gyrus (x = −24, y = 48, z = −8). The colour bar indicates z-values.
The region of interest analysis showed that the patients with Parkinson’s disease with hypersexuality had stronger activity during exposure to sexual compared with other rewards visual cues in the OFF medication states in the following regions: orbitofrontal cortex (left: P < 0.001, right: P < 0.001), anterior cingulate cortex (Left: P < 0.001, Right: P < 0.001), posterior cingulate cortex (left: P < 0.001, right: P < 0.001), ventral striatum (left: P < 0.001, right: P < 0.001) and hypothalamus (left: P < 0.001, right: P < 0.001). Similar activations were observed during the ON medication state, and there were no significant differences in activations between OFF and ON scans.
Whole-brain functional magnetic resonance imaging analysis
Common brain activity between patients with Parkinson’s disease with hypersexuality and Parkinson’s disease control patients
Patients with Parkinson’s disease with hypersexuality and Parkinson’s disease control patients showed similar significant BOLD signal increases during exposure to sexual versus neutral visual cues in both ON and OFF medication states bilaterally in middle temporal gyrus and middle occipital gyrus (Fig. 1A and B; Supplementary Tables 2A and C, Supplementary DataA and C). For the same contrast in the OFF state, patients with Parkinson’s disease with hypersexuality and Parkinson’s disease control patients showed similar significant BOLD signal decreases bilaterally in isthmus of cingulate gyrus [Brodmann area (BA) 29 and 30], parahippocampal gyrus and cuneus (BA 17) (Fig. 2A and B; Supplementary Tables 2B and D, Supplementary DataB and D).
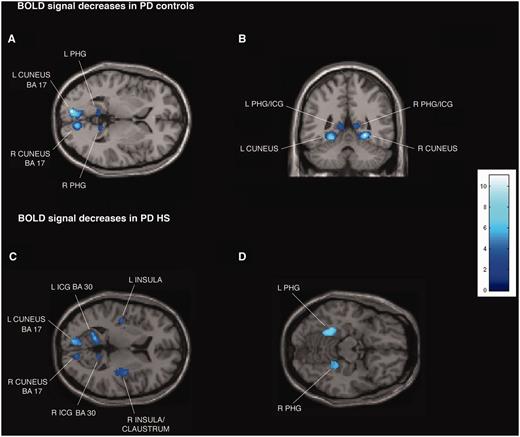
{kind=link}
Transverse and coronal sections of statistical parametric maps showing significant BOLD signal decreases (dark and light blue areas) associated with exposure to common sexual imagery in 12 Parkinson’s disease (PD) control patients (A and B) and in 12 patients with Parkinson’s disease with hypersexuality (HS) (C and D) during the OFF medication state. BOLD signal decreases are illustrated in (A and B) left and right parahippocampal gyrus (PHG), isthmus of cingulate gyrus (ICG) and cuneus (x = −9, y = −47, z = 2), (C) left and right parahippocampal gyrus, cuneus, isthmus of cingulate gyrus, insula and right claustrum (x = −42, y = 14, z = 8), and (D) left and right parahippocampal gyrus (x = 22, y = 38, z = −14). In the ON medication state, in the group of 12 patients with Parkinson’s disease with hypersexuality, no BOLD signal decreases are detected. The colour bar indicates z-values.
Brain activity specific to patients with Parkinson’s disease with hypersexuality
Whole-brain analysis confirmed the region of interest results and showed additional significant BOLD signal increases in the group of patients with Parkinson’s disease with hypersexuality compared with Parkinson’s disease control patients during exposure to sexual versus neutral visual cues in the OFF and ON medication states in the following regions: bilateral anterior prefrontal cortex (BA 10) and superior parietal lobule (BA 5 and 7), and right lateralized activation in the inferior parietal lobule (BA 40) (Fig. 1C–H; Supplementary Tables 2A and C, Supplementary DataA and C). For the same contrast in the OFF state, patients with Parkinson’s disease with hypersexuality compared with Parkinson’s disease control patients showed significant BOLD signal decreases bilaterally in insula and right claustrum (Fig. 2C and D; Supplementary Tables 2B and D, Supplementary DataB and D). There were no different BOLD signal increases during exposure to sexual versus neutral visual cues in patients with Parkinson’s disease with hypersexuality between the OFF and ON medication states.
Whole-brain analysis also confirmed the region of interest results in patients with Parkinson’s disease with hypersexuality when compared sexual versus other rewards visual cues and showed additional significant BOLD signal increases in the OFF and ON medication states in bilateral dorsolateral prefrontal cortex (BA 9), and right lateralized activation in anterior prefrontal cortex (BA 10) and parahippocampal gyrus.
Effect of l-DOPA medication in brain activity in patients with Parkinson’s disease with hypersexuality
In the group of patients with Parkinson’s disease with hypersexuality in the ON medication state, the decreases observed in the OFF state and in the OFF and ON state in Parkinson’s disease control patients (reduced BOLD signal during exposure to sexual versus neutral visual cues in isthmus of cingulate gyrus, parahippocampal gyrus, cuneus, insula and claustrum) were eliminated (Supplementary Tables 2B and D, Supplementary DataB and D).
Brain activity related to other reward visual cues in patients with Parkinson’s disease with hypersexuality
We found no significant within or between group differences for patients with Parkinson’s disease with hypersexuality and Parkinson’s disease control patients in any other reward condition minus baseline (neutral pictures) (e.g. dopaminergic drug cues versus neutral) in either ON and OFF scans.
Correlations
In the group of patients with Parkinson’s disease with hypersexuality, sexual desire post-exposure to sexual visual cues correlated in the OFF medication state, with the activity contrast (sex versus neutral) in the posterior cingulate cortex (r = 0.78, P < 0.01) and ventral striatum (r = 0.80, P < 0.01) (Fig. 3A and B), and in the ON medication state with the activity in the anterior cingulate cortex (r = 0.87, P < 0.001) and medial orbitofrontal cortex (r = 0.65, P < 0.05) (Fig. 3C and D). No correlations were found in the patients with Parkinson’s disease with hypersexuality group between ‘liking’ the sexual cues and brain activity during exposure to sexual versus neutral visual cues in both the ON and OFF medication states. No correlations were found between desire scores and activity contrasts in the the Parkinson’s disease control group in both the ON and OFF medication states (P > 0.1 in all cases).
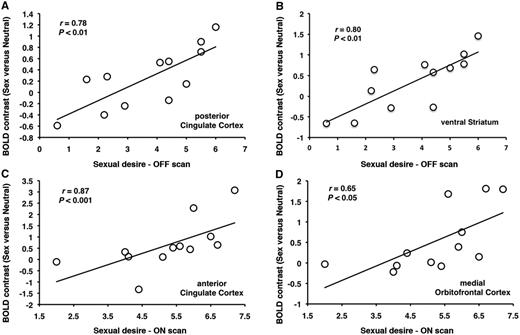
{kind=link}
In patients with Parkinson’s disease with hypersexuality, sexual desire post-exposure to sexual imagery correlates with increases in activation in (A) posterior cingulate cortex and (B) ventral striatum in the OFF medication state and with increases in activation in (C) anterior cingulate cortex and (B) and medial orbitofrontal cortex during the ON medication state.
Discussion
Hypersexuality is a relatively common unwanted effect of dopaminergic treatment in Parkinson’s disease, which can occasionally result in devastating social consequences including divorce, ruination of a person’s reputation and even arrest. Our study aimed to explore the mechanisms underpinning dopaminergic drug linked hypersexuality in patients with Parkinson’s disease. We have used behavioural assessments of sexual motivation and used a functional MRI paradigm with common portrayals of sexuality (similar to those projected by mass media) as visual cues, and we confirmed that exposure to such cues was adequate to increase the sexual desire of patients with Parkinson’s disease with hypersexuality by activating brain regions linked to sexual motivation (see Table 4 for summary of results).
|
|
|
|||
|---|---|---|---|---|
Summary of results showing differences and commonalities in regional BOLD signal increases and decreases in patients with Parkinson’s disease with and without hypersexuality in the ON and OFF medication states
| Parkinson’s disease with hypersexuality
|
Parkinson’s disease control patients
|
|||
|---|---|---|---|---|
| OFF and ON increases | OFF decreases | ON decreases | OFF and ON increases | OFF and ON decreases |
| Middle temporal gyrus | Isthmus of cingulate gyrus | Middle temporal gyrus | Isthmus of cingulate gyrus | |
| Middle occipital gyrus | Parahippocampal gyrus | Middle occipital gyrus | Parahippocampal gyrus | |
| Cuneus | Cuneus | |||
| Ventral striatum | Claustrum | |||
| Amygdala | Insula | |||
| Hypothalamus | ||||
| Anterior cingulate cortex | ||||
| Posterior cingulate cortex | ||||
| Orbitofrontal cortex | ||||
| Anterior prefrontal cortex | ||||
| Superior parietal lobule | ||||
| Inferior parietal lobule | ||||
In agreement with functional imaging studies of visual sexual stimulation in healthy individuals (Redouté et al., 2000; Arnow et al., 2002; Walter et al., 2008), exposure to visual sexual cues activated parieto-temporal-occipital visual processing regions such as the middle occipital gyrus and middle temporal gyrus in the groups of patients with Parkinson’s disease with hypersexuality and the Parkinson’s disease control patients. In patients with Parkinson’s disease with hypersexuality, exposure to sexual visual cues in both the ON and OFF medication conditions showed increased activation in the orbitofrontal cortex, anterior cingulate cortex, posterior cingulate cortex, ventral striatum, anterior prefrontal cortex, superior parietal lobule, amygdala and the hypothalamus, compared with the Parkinson’s disease control patients. Patients with Parkinson’s disease with hypersexuality showed significantly increased sexual desire and liking of the sexual content following exposure to sexual cues. Their sexual desire was increased more when they were ON l-DOPA compared with OFF medication after exposure to sexual content. The crucial role of the anterior cingulate cortex, posterior cingulate cortex, ventral striatum and orbitofrontal cortex in sexual motivation was underlined by the fact that increased activation in these regions correlated with increased sexual desire. However, no correlation was found between regional brain activity and ‘liking’ scores.
Animal studies have shown that dopamine is involved in sexual motivation and urge triggered by sexual cues in areas including the ventral striatum, hypothalamus, medial prefrontal cortex, amygdala and anterior cingulate cortex, and that a sensitizing regime of dopaminergic drugs can lead to exaggerated sexual pursuit and predation (Fiorino and Phillips, 1999; Nocjar and Panksepp, 2002; Afonso et al., 2009; Pfaus, 2010; Stolzenberg and Numan, 2011). Concurrent experience with dopaminergic drugs and sexual activity is required for development of compulsive sexual behaviour (Frohmader et al., 2011), mirroring the delayed onset of hypersexuality in Parkinson’s disease following treatment with dopaminergic drugs (Giladi et al., 2007). Moreover, recent work has shown that drugs of abuse can activate the same neuronal systems as a sexual reward, with overlap occurring in regions including the hypothalamus and anterior cingulate cortex (Frohmader et al., 2010). Our findings taken together with previous animal work support the incentive sensitization theory as a framework for understanding the mechanisms of dopaminergic drug induced hypersexuality in Parkinson’s disease. The incentive sensitization theory holds that dopamine motivates the pursuit of rewards by attributing incentive salience to reward-related stimuli (such as sexual visual cues), triggering pursuit (‘wanting’) of sex and in the case of hypersexuality, sex cues may be attributed with pathological incentive salience (Berridge et al., 2009). Incentive sensitization theory further holds that ‘wanting’ for rewards, mediated by ventral striatum dopamine-related neurocircuitry, may grow over time independently of reward ‘liking’ as an individual develops compulsive reward seeking (Berridge et al., 2009), and it is notable here that dopamine increased cue-evoked sexual desire (‘wanting’), but not rated ‘liking’ of the sexual images in patients with Parkinson’s disease with hypersexuality.
As the majority (8 of 12) of patients with Parkinson’s disease with hypersexuality had at least one additional impulse control disorder (e.g. pathological gambling, binge eating, etc.), we wanted to test whether the changes in behavioural assessments and brain activation following reward cue exposure were specific to sex or extended to other rewards. Self-reported desire and liking for gambling, drugs and food were not different within or between the groups of patients with Parkinson’s disease with hypersexuality and Parkinson’s disease control patients before and after the ON and OFF scans. Similarly, we did not observe any differences between patients with Parkinson’s disease with hypersexuality and Parkinson’s disease control patients in neural response to other rewarding visual cues in the functional MRI analysis indicating that our imaging results were also specific to hypersexuality. However, we wanted also to explore the specific brain activity related to sexual visual cues compared with other rewards. The findings revealed strong activation in both the ON and OFF state scans in the orbitofrontal cortex, anterior cingulate cortex, posterior cingulate cortex, ventral striatum, anterior prefrontal cortex, dorsolateral prefrontal cortex, parahippocampal gyrus and hypothalamus, confirming the specific importance of these regions in sexual motivation and desire, at least in the context of hypersexuality.
This selective enhancement for sexual cues is notable. Work in animal models by Berridge and colleagues (Mahler and Berridge, 2009, 2012; DiFeliceantonio and Berridge, 2012) has shown that stimulating opioid circuitry in the amygdala and striatum can focus cue-triggered ‘wanting’ very narrowly in a winner-take-all fashion. Under such conditions, a single favourite reward cue becomes a highly potent ‘motivational magnet’ that pulls all attraction to itself, at the expense of other reward cues, even in animals that were also attracted to those other rewards (Mahler and Berridge, 2009, 2012; DiFeliceantonio and Berridge, 2012). Similarly, the sexual reward cues were associated with enhanced amygdala activity relative to other reward cues in individuals with hypersexuality and may have been selectively targeted with enhanced incentive salience in a ‘winner-take-all manner’, even in those individuals with co-morbid impulse control disorders.
The increased activation in the ventral striatum could be related to increased dopamine release in the ventral striatum (Schott et al., 2008), consistent with PET data (O’Sullivan et al., 2011) and previous studies showing increased dopamine release related to sexual cues in the ventral striatum of sensitized animals (Fiorino and Phillips, 1999). Lesions in the hypothalamus and amygdala are known to impair sexual motivation in male and female rodents, and hypothalamic and amygdala activation tracks sexual arousal in functional MRI studies (Hamann et al., 2004). The hypothalamus has been shown to play a central role in sexual behaviour and is believed to be involved in the autonomic component of sexual arousal (Allen et al., 1989; Kupfermann et al., 1991; Meisel and Sachs, 1994; Georgiadis et al., 2010).
The anterior cingulate cortex is another paralimbic area connected to forebrain regions and controlling a number of autonomic and neuroendocrine functions (Stoléru et al., 1999). Previous studies have connected the activation of the anterior cingulate cortex with the degree of penile tumescence (Redouté et al., 2000) and with the motivational component of sexual arousal and desire (Redouté et al., 2000; Arnow et al., 2002; Karama et al., 2002; Walter et al., 2008). Parietal areas and the posterior cingulate cortex have also been linked to cue specificity in reward desire (Garavan et al., 2000). The orbitofrontal cortex has been linked to explicit ratings of anticipated pleasure and desire, and it is believed to have a role in mediating subjective reward experience (Kringelbach, 2005). Furthermore, activation of the orbitofrontal cortex and prefrontal cortex has been suggested to be associated with a pleasant bodily sensation being internally represented during sexual arousal (Stoléru et al., 2003). We found that rated sexual desire was correlated with activity in cortical regions (including anterior cingulate cortex and orbitofrontal cortex) in addition to ventral striatum, which may be consistent with the notion that incentive salience ‘wanting’ is not directly experienced in conscious awareness, and that explicit, conscious ‘wanting’ requires additional cortical processing of ventral striatum outputs (Berridge and Robinson, 1995).
In our study, enhanced brain activity after exposure to sexual cues was not influenced by the administration of l-DOPA as suggested by the similar BOLD signal increases in the ON and OFF medication scans. However, the absence of an acute l-DOPA effect does not eliminate the effect of synergistic interaction of mesolimbic dopamine state and the presence of sexual cues in stimulating enhanced ‘wanting’ for sexual reward in the group of patients with Parkinson’s disease with hypersexuality (Berridge, 2012; Oei et al., 2012). The patients with Parkinson’s disease with hypersexuality, even in the OFF state are not entirely drug-free and are presumably sensitized, and in animal studies, although acute amphetamine and sensitization effects can be additive (Tindell et al., 2005), sensitization caused by previous amphetamine administration causes reward cues to trigger excessive pursuit of their associated reward, even when sensitized rats are tested in a drug-free state (Wyvell and Berridge, 2001).
Interestingly, in the group of patients with Parkinson’s disease with hypersexuality after acute l-DOPA challenge, the decreases observed when OFF their drugs and in the OFF and ON state in Parkinson’s disease control patients (reduced BOLD signal during exposure to sexual versus neutral visual cues in isthmus of cingulate gyrus, parahippocampal gyrus, cuneus, insula and claustrum) were eliminated (for a similar effect of dopaminergic drug challenge in cocaine addiction, see Volkow et al., 2010). These results suggest that dopamine drugs could eliminate the deactivation of brain regions, which may be related to plastic changes in an effort to inhibit hypersexuality stimulation. Although it is difficult to translate excitation or inhibition of neuronal firing into BOLD signal increases or decreases in activated voxels (Georgopoulos et al., 1982; Batini et al., 1984), dopamine drugs could release this inhibition through the activity of inhibitory interneurons acting within local neuronal circuits in the cerebral cortex. The release of neuronal inhibition in the isthmus of cingulate gyrus, parahippocampal gyrus, cuneus, insula and claustrum when patients are ON their dopaminergic drugs could be associated with concurrent behavioural changes as in this study was an associated increase in sexual desire scores. This theory is supported by the complete cessation of hypersexuality after withdrawal from dopamine receptor agonist medications (Mamikonyan et al., 2008; Munhoz et al., 2009).
Inhibition of deactivation of insula through dopamine drugs could lead to the expression of pathological sexual behaviour, as this paralimbic area forms connections to regions involved in autonomic regulation (Oppenheimer et al., 1992), relates highly processed sensory information with motivational states (Stoléru et al., 1999) and, in combination with the secondary somatosensory cortex, has been reported to be involved in the perceived urge to express overt sexual behaviour (Mouras et al., 2003). The claustrum has also been shown to be involved in emotional and motivational responses in animals (Hamamura et al., 1997) and humans (Reiman et al., 1989) and is thought to be associated with sexual motivation (Rees et al., 2007). Deactivation in parahippocampal regions has been reported following presentation of sexually explicit video clips; however, the potential role of these deactivations is not well understood (Redouté et al., 2000).
In conclusion, this is the first study to examine the mechanisms underpinning hypersexuality in Parkinson’s disease, and the findings lend support to an account based on exaggerated cue-triggered incentive salience-based motivation. Further, our findings have some implications with respect to the potential of cue-exposure via access to mass media to influence pathological sexual behaviour in Parkinson’s disease. Stimulation by common erotic cues of individuals prone to hypersexuality is able to provide a motivational impetus for seeking sexual reward through increased and decreased activations of cerebral cortex, which could potentially lead to social and psychologically harmful consequences. Application of restrictions on type and quantity of exposure to erotic visual cues might therefore limit the onset of abnormal excessive or deviant sexual behaviour in vulnerable populations such as those receiving dopaminergic drugs.
Funding
This work was supported by funding from Parkinson’s UK (J-0704).
Supplementary material
Supplementary material is available at Brain online.